4.2 – Ion and Water Balance on Land
 |
4.4. Explain how ion and water balance function in sample animal terrestrial systems. |
The content of this chapter was adapted from the Concepts of Biology-1st Canadian Edition open textbook by Charles Molnar and Jane Gair (Chapter 22 – Kidneys and Osmoregulation).
Although the kidneys are the major osmoregulatory organ, the skin and lungs also play a role in the process. Water and electrolytes are lost through sweat glands in the skin, which helps moisturize and cool the skin surface, while the lungs expel a small amount of water in the form of mucous secretions and via evaporation of water vapor.
Kidneys: the main osmoregulatory organ
The kidneys, illustrated in Figure 4.6, are a pair of bean-shaped structures that are located just below and posterior to the liver in the peritoneal cavity. The adrenal glands sit on top of each kidney and are also called the suprarenal glands. Kidneys filter blood and purify it. All the blood in the human body is filtered many times a day by the kidneys; these organs use up almost 25 percent of the oxygen absorbed through the lungs to perform this function. Oxygen allows the kidney cells to efficiently manufacture chemical energy in the form of ATP through aerobic respiration. The filtrate coming out of the kidneys is called urine.
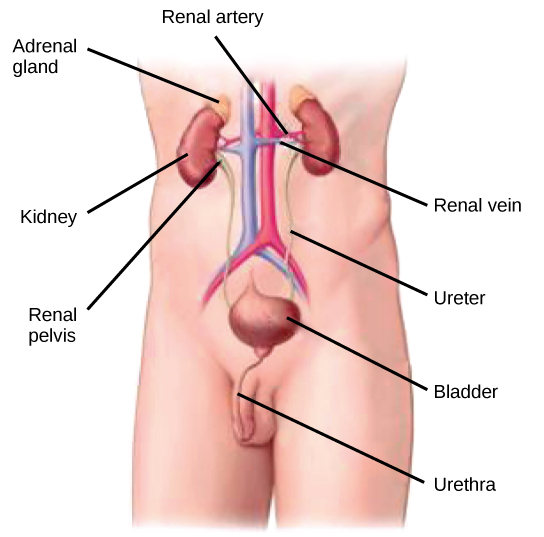
Figure 4.6. Kidneys filter the blood, producing urine that is stored in the bladder prior to elimination through the urethra. (credit: modification of work by NCI)
Kidney structure
Externally, the kidneys are surrounded by three layers, illustrated in Figure 4.7. The outermost layer is a tough connective tissue layer called the renal fascia. The second layer is called the perirenal fat capsule, which helps anchor the kidneys in place. The third and innermost layer is the renal capsule. Internally, the kidney has three regions—an outer cortex, a medulla in the middle, and the renal pelvis in the region called the hilum of the kidney. The hilum is the concave part of the bean-shape where blood vessels and nerves enter and exit the kidney; it is also the point of exit for the ureters. The renal cortex is granular due to the presence of nephrons—the functional unit of the kidney. The medulla consists of multiple pyramidal tissue masses, called the renal pyramids. In between the pyramids are spaces called renal columns through which the blood vessels pass. The tips of the pyramids, called renal papillae, point toward the renal pelvis. There are, on average, eight renal pyramids in each kidney. The renal pyramids along with the adjoining cortical region are called the lobes of the kidney. The renal pelvis leads to the ureter on the outside of the kidney. On the inside of the kidney, the renal pelvis branches out into two or three extensions called the major calyces, which further branch into the minor calyces. The ureters are urine-bearing tubes that exit the kidney and empty into the urinary bladder.
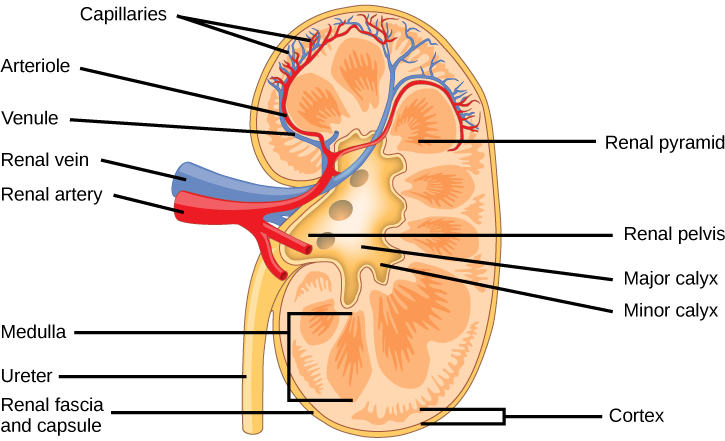
Figure 4.7 The internal structure of the kidney is shown. (credit: modification of work by NCI)
 |
Question 4.4
Which of the following statements about the kidney is false? |
Because the kidney filters blood, its network of blood vessels is an important component of its structure and function. The arteries, veins, and nerves that supply the kidney enter and exit at the renal hilum. Renal blood supply starts with the branching of the aorta into the renal arteries(which are each named based on the region of the kidney they pass through) and ends with the exiting of the renal veins to join the inferior vena cava. The renal arteries split into several segmental arteries upon entering the kidneys. Each segmental artery splits further into several interlobar arteries and enters the renal columns, which supply the renal lobes. The interlobar arteries split at the junction of the renal cortex and medulla to form the arcuate arteries. The arcuate “bow-shaped” arteries form arcs along the base of the medullary pyramids. Corticalradiate arteries, as the name suggests, radiate out from the arcuate arteries. The cortical radiate arteries branch into numerous afferent arterioles and then enter the capillaries supplying the nephrons. Veins trace the path of the arteries and have similar names, except there are no segmental veins.
As mentioned previously, the functional unit of the kidney is the nephron, illustrated in Figure 4.8. Each kidney is made up of over one million nephrons that dot the renal cortex, giving it a granular appearance when sectioned sagittally. There are two types of nephrons— cortical nephrons (85 percent), which are deep in the renal cortex, and juxtamedullary nephrons, which lie in the renal cortex close to the renal medulla. A nephron consists of three parts—a renal corpuscle, a renal tubule, and the associated capillary network, which originates from the cortical radiate arteries.
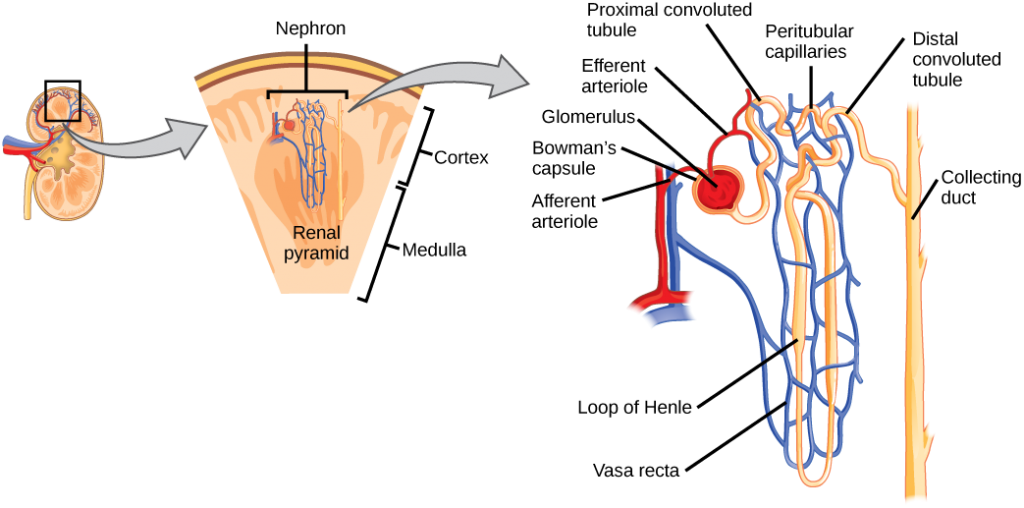
Figure 4.8. The nephron is the functional unit of the kidney. The glomerulus and convoluted tubules are located in the kidney cortex while collecting ducts are located in the pyramids of the medulla. (credit: modification of work by NIDDK)
Nephrons: the functional unit
Nephrons take a simple filtrate of the blood and modify it into the urine. Many changes take place in the different parts of the nephron before the urine is created for disposal. The term forming urine will be used hereafter to describe the filtrate as it is modified into the true urine. The principal task of the nephron population is to balance the plasma to homeostatic set points and excrete potential toxins in the urine. They do this by accomplishing three principle functions—filtration, reabsorption, and secretion. They also have additional secondary functions that exert control in three areas: blood pressure (via production of renin), red blood cell production (via the hormone EPO), and calcium absorption (via conversion of calcidiol into calcitriol, the active form of vitamin D).
 |
Go to this website to see another coronal section of the kidney and to explore an animation of the workings of nephrons. |
|
Question 4.5
Which of the following statements about the nephron is false? |
Kidney function and physiology
Kidneys filter blood in a three-step process. First, the nephrons filter blood that runs through the capillary network in the glomerulus. Almost all solutes, except for proteins, are filtered out into the glomerulus by a process called glomerular filtration. Second, the filtrate is collected in the renal tubules. Most of the solutes get reabsorbed in the PCT by a process called tubular reabsorption. In the loop of Henle, the filtrate continues to exchange solutes and water with the renal medulla and the peritubular capillary network. Water is also reabsorbed during this step. Then, additional solutes and wastes are secreted into the kidney tubules during tubular secretion, which is, in essence, the opposite process of tubular reabsorption. The collecting ducts collect filtrate coming from the nephrons and fuse in the medullary papillae. From here, the papillae deliver the filtrate, now called urine, into the minor calyces that eventually connect to the ureters through the renal pelvis. This entire process is illustrated in Figure 4.9.
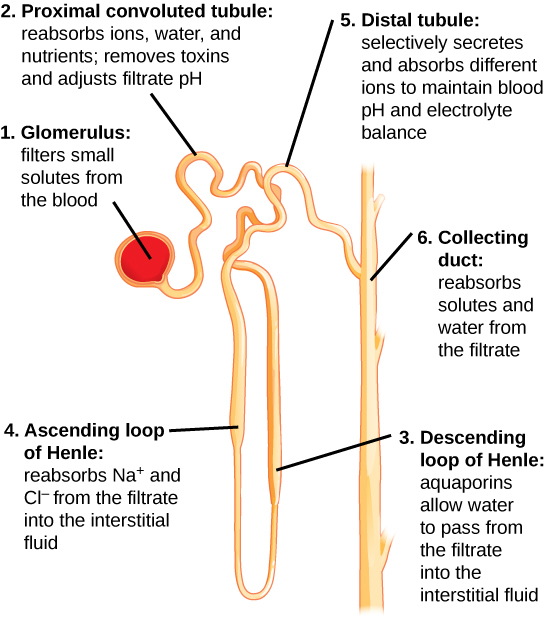
Figure 4.9. Each part of the nephron performs a different function in filtering waste and maintaining homeostatic balance. (1) The glomerulus forces small solutes out of the blood by pressure. (2) The proximal convoluted tubule reabsorbs ions, water, and nutrients from the filtrate into the interstitial fluid, and actively transports toxins and drugs from the interstitial fluid into the filtrate. The proximal convoluted tubule also adjusts blood pH by selectively secreting ammonia (NH3) into the filtrate, where it reacts with H+ to form NH4+. The more acidic the filtrate, the more ammonia is secreted. (3) The descending loop of Henle is lined with cells containing aquaporins that allow water to pass from the filtrate into the interstitial fluid. (4) In the thin part of the ascending loop of Henle, Na+ and Cl- ions diffuse into the interstitial fluid. In the thick part, these same ions are actively transported into the interstitial fluid. Because salt but not water is lost, the filtrate becomes more dilute as it travels up the limb. (5) In the distal convoluted tubule, K+ and H+ ions are selectively secreted into the filtrate, while Na+, Cl-, and HCO3- ions are reabsorbed to maintain pH and electrolyte balance in the blood. (6) The collecting duct reabsorbs solutes and water from the filtrate, forming dilute urine. (credit: modification of work by NIDDK)
Tubular reabsorption and secretion
Tubular reabsorption occurs in the PCT part of the renal tubule. Almost all nutrients are reabsorbed, and this occurs either by passive or active transport. Reabsorption of water and some key electrolytes are regulated and can be influenced by hormones. Sodium (Na+) is the most abundant ion and most of it is reabsorbed by active transport and then transported to the peritubular capillaries. Because Na+ is actively transported out of the tubule, water follows it to even out the osmotic pressure. Water is also independently reabsorbed into the peritubular capillaries due to the presence of aquaporins, or water channels, in the PCT. This occurs due to the low blood pressure and high osmotic pressure in the peritubular capillaries. However, every solute has a transport maximum and the excess is not reabsorbed.
In the loop of Henle, the permeability of the membrane changes. The descending limb is permeable to water, not solutes; the opposite is true for the ascending limb. Additionally, the loop of Henle invades the renal medulla, which is naturally high in salt concentration and tends to absorb water from the renal tubule and concentrate the filtrate. The osmotic gradient increases as it moves deeper into the medulla. Because two sides of the loop of Henle perform opposing functions, as illustrated in Figure 4.10, it acts as a countercurrent multiplier. The vasa recta around it acts as the countercurrent exchanger.
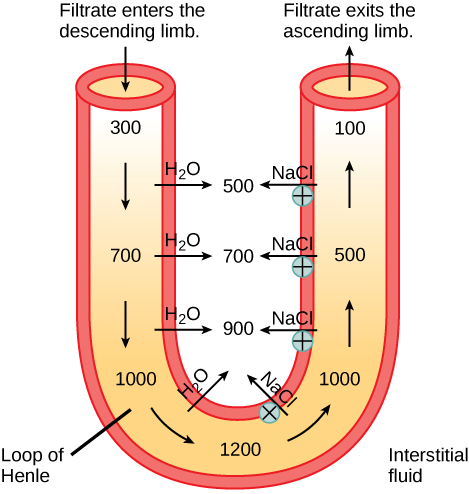
Figure 4.10. The loop of Henle acts as a countercurrent multiplier that uses energy to create concentration gradients. The descending limb is water permeable. Water flows from the filtrate to the interstitial fluid, so osmolality inside the limb increases as it descends into the renal medulla. At the bottom, the osmolality is higher inside the loop than in the interstitial fluid. Thus, as filtrate enters the ascending limb, Na+ and Cl- ions exit through ion channels present in the cell membrane. Further up, Na+ is actively transported out of the filtrate and Cl- follows. Osmolarity is given in units of milliosmoles per liter (mOsm/L).
By the time the filtrate reaches the DCT, most of the urine and solutes have been reabsorbed. If the body requires additional water, all of it can be reabsorbed at this point. Further reabsorption is controlled by hormones, which will be discussed in a later section. Excretion of wastes occurs due to lack of reabsorption combined with tubular secretion. Undesirable products like metabolic wastes, urea, uric acid, and certain drugs, are excreted by tubular secretion. Most of the tubular secretion happens in the DCT, but some occurs in the early part of the collecting duct. Kidneys also maintain an acid-base balance by secreting excess H+ ions.
Although parts of the renal tubules are named proximal and distal, in a cross-section of the kidney, the tubules are placed close together and in contact with each other and the glomerulus. This allows for the exchange of chemical messengers between the different cell types. For example, the DCT ascending limb of the loop of Henle has masses of cells called macula densa, which are in contact with cells of the afferent arterioles called juxtaglomerular cells. Together, the macula densa and juxtaglomerular cells form the juxtaglomerular complex (JGC). The JGC is an endocrine structure that secretes the enzyme renin and the hormone erythropoietin. When hormones trigger the macula densa cells in the DCT due to variations in blood volume, blood pressure, or electrolyte balance, these cells can immediately communicate the problem to the capillaries in the afferent and efferent arterioles, which can constrict or relax to change the glomerular filtration rate of the kidneys.
|
To learn more about the vascular system of kidneys, click through this review and the steps of blood flow. |
